p53 Informationp53 informationp53 story
p53 monoclonal antibodiesp53 pathwaysp53 genep53 protein(s)p63/p73 protein(s)p53 evolutionp53 polymorphismp53 and cancer
|
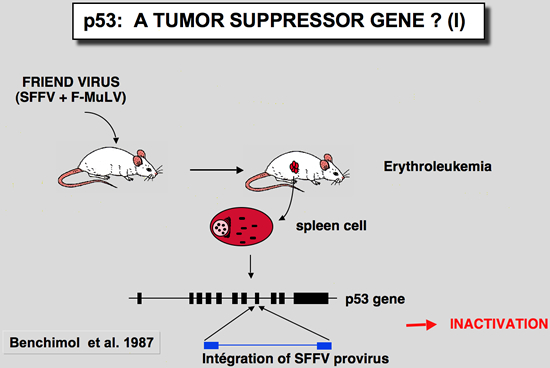
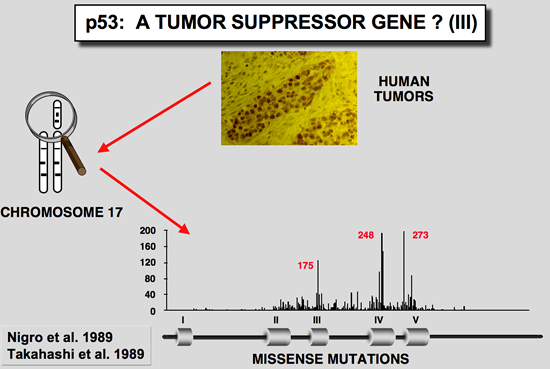
p53 as a Tumour suppressorp53 in Friend murine erythroleukemiaIn these tumors induced by the Friend virus, the p53 gene found in the tumor cells is very often rearranged, leading to an absence of expression or the synthesis of a truncated or mutant protein (Mowat et al. 1985) The mutation often affects one of the conserved blocks of the protein (Munroe et al. 1988). In all cases studied, the second allele is either lost through loss of the chromosome, or inactived by deletion. In this tumor model, functional inactivation of the p53 gene seems to confer a selective growth advantage to erythroid cells during the development of Friend leukemia in vivo.
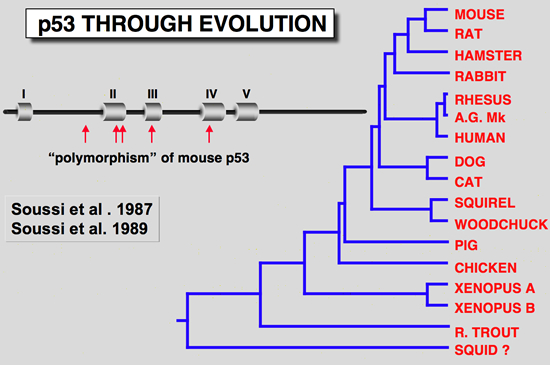
There are mutations in the various murine p53 cDNA clonesThe finding that one murine p53 cDNA clone isolated from the F9 cell failed to cooperate with an activated Ha-ras gene was another clue that the p53 cDNA clones differ from one another in their behavior (Finlay et al. 1988). Examination of all murine p53 cDNA clones available revealed several codon changes which were primarily assumed to be due to polymorphism. However, comparison of these sequence differences with p53 from lower species indicated that some of them occur in highly conserved regions (Soussi et al. 1990; Soussi et al. 1987), a feature which is not linked to polymorphism. Careful reinvestigation of all sequences led to the conclusion that the F9 cDNA clone was a wild type, while most of the others used in transfection experiments contain point mutations which activate their transforming properties.
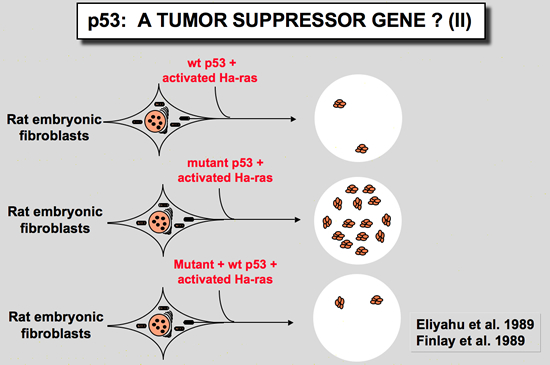
Wild type p53 has antiproliferative properties and does not cooperate with Ha-rasA new set of experiments has shown that cotransfection of a plasmid encoding wild type p53 reduced the transformation potential of plasmids encoding p53 and an activated Ha-ras gene (Eliyahu et al. 1989; Finlay et al. 1989). Furthermore, wild type p53 was shown to suppress transformation by a mixture of E1A or myc and an activated Ha-ras gene. These transformation experiments indicate that wild type p53 is a suppressor of cell transformation in vitro. p53 gene is mutated in a wide variety of human cancer
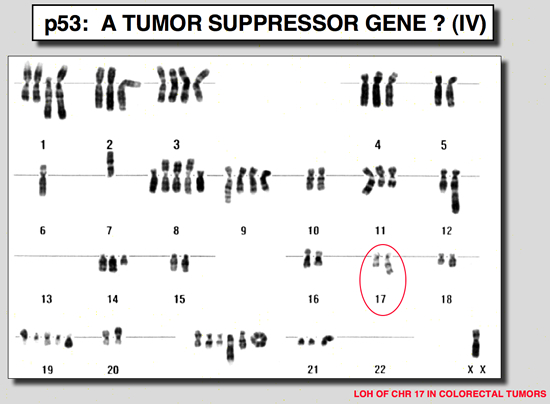
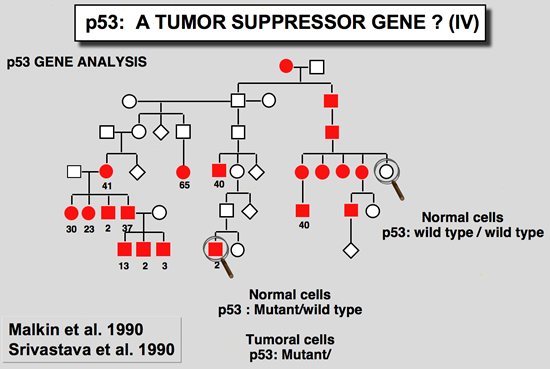
Germline mutation of the p53 gene are found in Li-Fraumeni patientsTransgenic mice carrying a mutant p53 gene develop many types of cancer, with a high proportion of sarcomas (Lavigueur et al. 1989). This observation led various authors to study patients with Li-Fraumeni syndrome. This syndrome presents as a familial association of a broad spectrum of cancers including osteosarcomas, breast cancer, soft tissue sarcoma and leukemias, appearing at a very early age. Statistical analysis predicts that 50 % of these individuals will have a tumor before the age of 30, and 90 % before the age of 70. Germ-line mutations in the p53 gene have been found in several families with this syndrome (Malkin et al. 1990; Srivastava et al. 1990). In all cases there is a strict correlation between transmission of the mutant allele and development of a cancer.
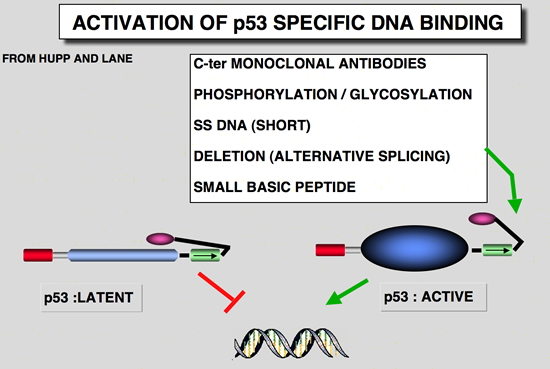
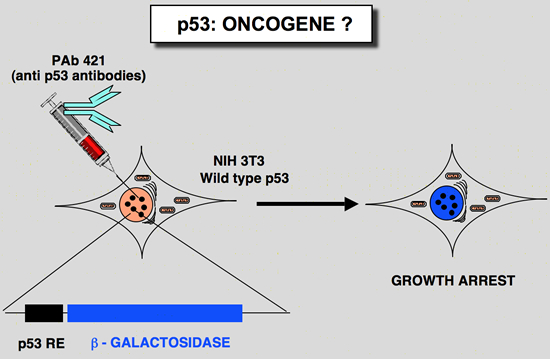
Why micro-injection of p53 monoclonal antibody induces a growth arrest ?The carboxy-terminus of Hp53 has been shown to play an important role in controlling the specific DNA binding function. Wild type p53 is found in a latent form that does not bind to DNA. The specific DNA binding activity was shown to be activated by various pathways: phosphorylation (Hupp et al., 1992), antibody specific for the carboxy-terminus of the protein (Hupp et al., 1992), small peptides which could mimic the carboxy-terminus of the p53 (Hupp et al., 1995), short single stranded DNA (Jayaraman & Prives, 1995), deletion of the last 30 amino-acids (Hupp et al., 1992) and the interaction with a cellular protein (Jayaraman et al., 1997). This observation suggest that micro-injection of antibodies such as PAb421 induces an activation of the transcriptional activity of p53. Such hypothesis have been confirmed (Hupp et al., 1995) Wild type p53 as a tumor suppressor gene and mutant p53 as a dominant oncogene ? Taken together, these data made it possible to define the p53 gene as a tumor suppressor gene. Yet unlike the Rb gene, which is the archetype of the tumor suppressor genes, the p53 gene has some original features. In particular, more than 95 % of alterations in the p53 gene are point mutations that produce a mutant protein, which in all cases has lost its transactivational activity (see above). Nevertheless, the synthesis of these mutant p53 proteins is not harmless for the cell. In paticular, it has been shown that some p53 mutants (depending on the site of mutation) exhibit a transdominant phenotype and are able to associate with wild-type p53 (expressed by the remaining wild-type allele) to induce the formation of an inactive heteroligomer (Milner and Medcalf 1991). Moreover, cotransfection of mutant p53 with an activated ras gene shows that some p53 mutants have high, dominant oncogenic activity (Halevy et al. 1990). These observations led to the proposal that several classes of mutant p53 exist, according to the site of mutation and its phenotype (Michalovitz et al. 1991): i) null mutations with totally inactive p53 that do not directly intervene in transformation; ii) dominant negative mutations with a totally inactive p53 that is still able to interfere with wild-type p53 expressed from the wild-type allele, and iii) positive dominant mutations where the normal function of p53 is altered but in this case the mutant p53 acquires an oncogenic activity that is directly involved in transformation. REFERENCES
|
Home | Our Work |p53 Info| p53 Database | p53 Link | Contact us |