p53 Information
p53 protein(s)p63/p73 protein(s)p53 evolutionp53 polymorphismp53 and cancer
|
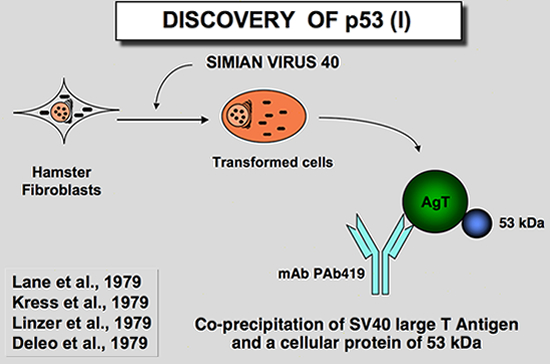
The Discovery of p53The discovery in 1979 of the p53 protein was the culmination of two types of studies involving a virologic approach and a serologic approach. The well known virologic approachStudies of SV40-transformed cells show that a 55-kDa protein is coprecipitated with the large-T antigen (Chang et al. 1979; Kress et al. 1979; Lane and Crawford 1979; Linzer and Levine 1979; Melero et l. 197). This association was shown to be the result of an in vivo association between the two proteins (Lane and Crawford 1979). It was then postulated that this protein could be encoded by the cellular genome. (It should be kept in mind that no middle-T was found for SV40 and that the molecular weight of this protein was similar to that of polyoma middle-T antigen). Linzer and Levine (Linzer and Levine 1979) found that the 54-kDa protein was overexpressed in a wide variety of murine SV40 transformed cells, but also in uninfected embryonic carcinoma cells. A partial peptide map from this 54-kDa protein was identical among the different cell lines, but was clearly different from the peptide map of SV40 large-T antigen (Kress et al. 1979; Linzer and Levine 1979). It was then postulated that SV40 infection or transformation of mouse cells stimulates the synthesis or stability of a cellular 54-kDa protein.
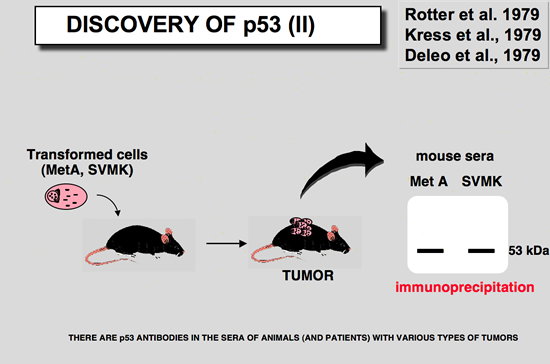
The serological approach, not so well knownIn 1979, DeLeo et al. (De Leo et al. 1979), showed that the humoral response of mice to some methylcholanthrene-induced tumor cell line such as MethA was directed toward the p53 protein. Later, it was found that animals bearing several types of tumors elicited an immune response specific for p53 (Kress et al. 1979; Melero et al. 1979; Rotter et al. 1980). In 1982, Crawford et al. (Crawford et al. 1982) first described antibodies against human p53 protein in 9% of breast cancer patient sera. No significant clinical correlation was reported, and at that time no information was available concerning mutations of the p53 gene. Caron de Fromentel et al. later found that such antibodies were present in sera of children with a wide variety of cancers (Caron de Fromentel et al. 1987). The average frequency was 12%, but the figure was 20 % in Burkitt Lymphoma.
REFERENCES
Chang C, Simmons DT, Martin MA and Mora PT (1979) Identification and partial characterization of new antigens from simian virus 40-transformed mouse cells. J Virol 31: 463-471. Crawford LV, Pim DC and Bulbrook RD (1982) Detection of antibodies against the cellular protein p53 in sera from patients with breast cancer. Int. J. Cancer 30: 403-408. De Leo AB, Jay G, Appella E, Dubois GC, Law LW and Old LJ (1979) Detection of a transformation-related antigen in chemically induced sarcomas and other transformed cells of the mouse. Proc Natl Acad Sci USA 76: 2420-2424. Kress M, May E, Cassingena R and May P (1979) Simian Virus 40-transformed cells express new species of proteins precipitable by anti-simian virus 40 serum. J. Virol. 31: 472-483. Lane DP and Crawford LV (1979) T antigen is bound to a host protein in SV40-transformed cells. Nature 278: 261-263. Linzer DIH and Levine AJ (1979) Characterization of a 54 K dalton cellular SV40 tumor antigen resent in SV40-transformed cells and in infected embryonal carcinoma cells. Cell 1: 43-52. Melero JA, Stitt DT, Mangel WF and Carroll RB (1979) Identification of new polypeptide species (48-55K) immunoprecipitable by antiserum to purified large T antigen and present in simian virus 40-infected and transformed cells. J. Virol. 93: 466-480. Rotter V, Witte ON, Coffman R and Baltimore D (1980) Abelson murine leukemia virus-induced tumors elicit antibodies against a host cell protein, p50. J Virol 36: 547-555. SOME REVIEWS (1983-1992)A must: The birth of p53 Crawford L (1983) The 53,000-dalton cellular protein and its role in transformation. Int. Rev. Exp. Path. 25: 1-50. |
Home | Our Work |p53 Info| p53 Database | p53 Link | Contact us |