p53 databaseImprove p53 mutation detection and report p53 mutation and cancer
|
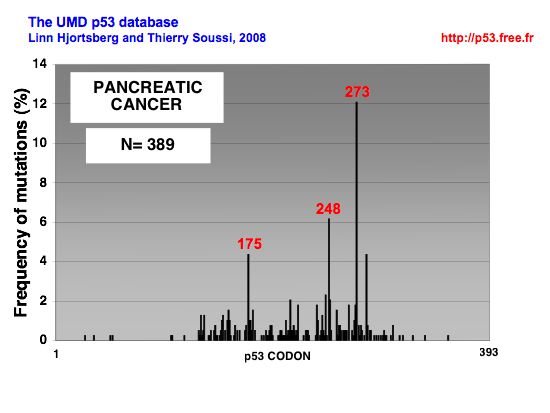
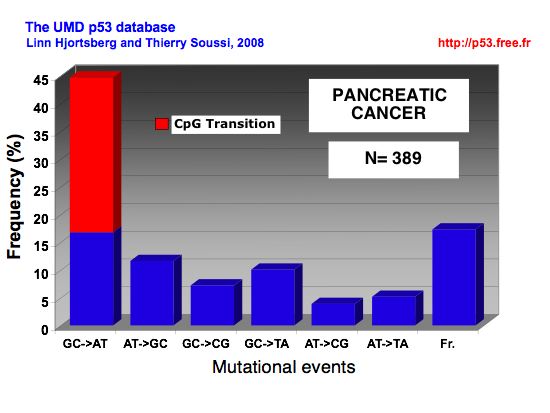
p53 MUTATIONS IN PANCREATIC CANCERPancreatic cancer Pancreatic cancer is associated with only a few known demographic and environmental risk factors and a handful of autosomal dominant genetic conditions. Multiple studies have established advanced age, smoking, and long-standing chronic pancreatitis as clear risk factors; diabetes and obesity also appear to confer increased risk. Increased risk has also been documented in relatives of PDAC patients, and it is estimated that 10% of PDAC cases are associated with an inherited predisposition based on familial clustering. Correspondingly, germline mutations have been linked to familial PDAC, including those targeting the tumor suppressor genes INK4A, BRCA2, and LKB1, the DNA mismatch repair gene MLH1 and the cationic trypsinogen gene PRSS1. BRCA1 mutation appears to confer increased susceptibility to PDAC, albeit with a lower associated risk than BRCA2. Given the low penetrance of PDAC and the typical age of onset associated with the above germline mutations, these genetic lesions appear to impact malignant progression of precursor lesions rather than cancer initiation. Supporting this hypothesis, INK4A and BRCA2 mutations are not detected in the earliest sporadic PDAC premalignant lesions but are only found in the later intermediate or advanced pancreatic intraepithelial neoplasm (PanIN) lesions. Additionally, mice engineered with germline INK4A mutations do not develop PDAC unless combined with activated K-RAS mutations (see below). The molecular analysis of evolving PDAC has provided a compendium of genetic lesions, often implicating known cancer genes and classical cancer signaling cascades. In many cases, these molecular events have been linked with defined histopathologic stages of PDAC progression Clinical and histopathologic studies have identified three PDAC precursor lesions: PanIN, mucinous cystic neoplasm (MCN), and intraductal papillary mucinous neoplasm (IPMN) . Of these precursor lesions, the most common and extensively studied is PanIN, which is found in the smaller-caliber pancreatic ducts. Consistent with a role in constraining malignant progression, p53 mutation appears in later-stage PanINs that have acquired significant features of dysplasia. In these more advanced PanINs, the selective pressure to eliminate p53 may stem in part from a collective accumulation of genetic damage, from telomere erosion and ROS, for example, resulting in the activation of p53-dependent DNA damage checkpoint responses. Thus, loss of p53 function could serve to enable the growth and survival of cells harboring procarcinogenic chromosomal aberrations. Given that human PDAC is characterized by profound aneuploidy and complex chromosomal rearrangements, as well as significant intratumoral genomic heterogeneity, a clear understanding of how p53 participates in genome stability mechanisms would provide important insights into disease pathogenesis and ultimately treatment. The rampant genomic instability in PDAC could serve to both fuel the rise of advanced disease and provide a basis for its resistance to therapeutic modalities. Finally, in many other cancer types, there exists a near reciprocal relationship in the loss of ARF and p53. This relationship likely reflects the fact that ARF inhibits MDM2-mediated targeting of the p53 protein for proteasomal degradation. Thus, ARF deficiency would result in marked reduction of p53 protein levels and attenuation of p53 pathway function in diverse cancer-relevant processes. On the other hand, in human PDAC, p53 mutations and ARF deletions coexist in 40% of cases, potentially pointing to nonoverlapping functions for these factors in pancreatic cancer suppression. Mounting evidence suggests that ARF possesses p53-independent functions including the inhibition of ribosomal RNA processing. In addition, ARF does not appear to neutralize the DNA damage checkpoint that would be activated upon genetic damage (e.g., induced by teleomere dysfunction), thereby necessitating the additional loss of p53 function as such signals intensify during PDAC progression. Alternatively, ARF deletion in PDAC could represent a "bystander" effect associated with mutational events targeting INK4A. The resolution of these issues will require systematic genetic analysis of various mutant genotype combinations in murine PDAC mouse models as well a greater understanding of the molecular actions of ARF versus p53 in PDAC tumor biology. Spectrum of p53 mutations in pancreatic cancer
|
Home | Our Work |p53 Info| p53 Database | p53 Link | Contact us |