| |
Our Work
The Yeast assay
p53 Chips and LDR
|
p53 Chips and LDR
Collaborative work:
Thierry Soussi (UPMC) and Francis Barany (Cornell)
 |
Pr F. Barany was awarded with the Mayent/Rotschild fellowship to come to the Institut Curie for our collaborative program
Francis barany: barany@mail.med.cornell.edu |
Cancers arise from the accumulation of multiple alterations in genes regulating cellular growth. Identification of such alterations is of importance as they can have clinical importance as they can be linked to the response to a particular therapy or to prognosis. Therefore, the detection of these alterations represents a significant challenge for the near futur.
The laboratory of Microbiology of Cornell University (New York) leaded by Pr Fr. Barany and the laboratoire de génotoxicologie des tumeurs (Institut Curie, Paris) have joined their effort to develop a platform for highthrouput detection of mutations in various genes involved in human neoplasia.
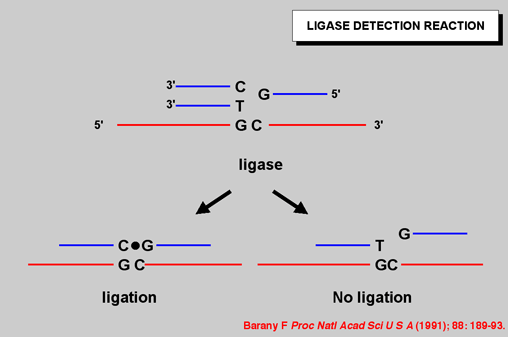
THE LIGASE DETECTION REACTION
|
Figure 1: The ligase detection reaction relies on the high specificity of DNA ligase to discriminate base variations in DNA (Barany, 1990; Barany, 1991a; Barany and Gelfand, 1991). A primary PCR reaction amplifies the region of DNA containing mutations or polymorphisms. Mutation specific discriminating oligonucleotides hybridize adjacent to a common primer, resulting in the formation of a short DNA duplex containing a nick between the oligonucleotides. If there is a perfect match at the nick, then the oligonucleotides will be covalently linked by DNA ligase to generate a uniquely labelled ligation product. A single base mismatch at the junction inhibits ligation, and thus single-base mutations or polymorphisms are distinguished (Barany, 1991a) |
|
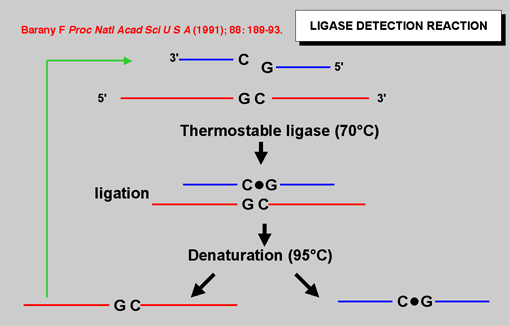
| Figure 2: Using thermostable DNA ligase dramatically improves the sensitivity of the reaction since ligation may be cycled to arithmetically increase the product |
|
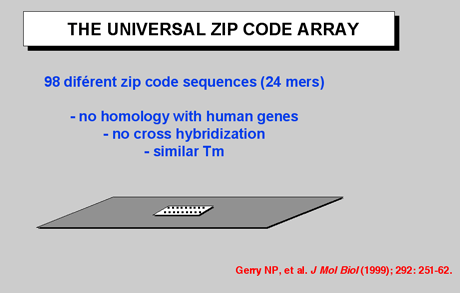
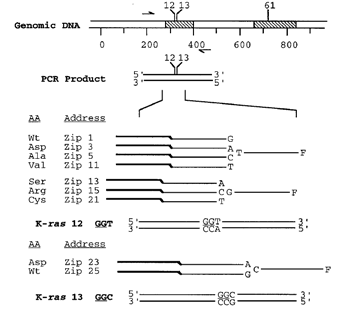
Figure 3: Currently, the "universal" array is designed with 64 “zip-code” addresses, but it may be increased to analyze as many as 4,000 separate bi-allelic polymorphisms. This approach relies on designing the 24-base zip-code sequences to be very different from each other and also to lack any sequence similarity to either the target sequence or to other sequences in the genome. In addition, divergent zip-code sequences are designed with similar thermodynamic properties. This allows for hybridizations to be carried out at a single temperature, 65 °C, resulting in a more stringent and rapid hybridization. In contrast, hybridization DNA microarrays require hybridizations at temperatures from 0 °C to 44 °C, since the immobilized probes on these arrays have a wide sequence variation and thus a range of Tm's. In addition, the wide sequence variation inherent in hybridization DNA microarrays results in differential hybridization efficiencies and is a source of increased background noise and false signals due to mismatch hybridization and nonspecific binding. In the universal DNA array format, these errors are significantly reduced, since the hybridization sequence is independent of the target gene. |
|
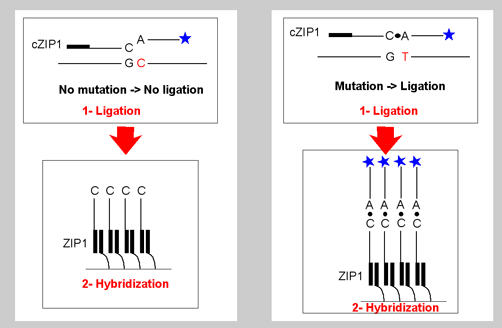
Figure 4: Each LDR product contains a fluorescent label on one end and a unique zip-code complement on the other end. The presence and type of each mutation is determined from the positions of those addresses at which labelled LDR products are captured. Since different mutation-specific LDR primers can be appended to the same set of zip-code complement sequences, this array is universal, and can be programmed to detect mutations in any gene. Each discriminating primer contains a 3’ base complementary to the mutation and a corresponding fluorescent label (currently Cy3, Cy5, and fluoresce in) at the 5’ end. The common primer contains a unique “zip-code” complement sequence at the 3’ end. In the presence of mutant target, adjacent primers are ligated together to form a product with both a fluorescent label and a zip-code complement. Thus, a given fluorescent signal captured at a unique address defines the type of mutation present in the clinical sample. In the example shown, a separate set of LDR primers targeted to a region distinct from the position of the mutation is used to quantify the total amount of that PCR fragment in the sample. Comparison of the ratio of mutant to specific fragment signal with known standards may be used to quantify low-abundance mutations |
.
|
|
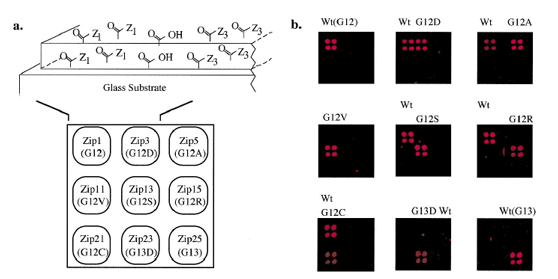
Figure 5 and 6:Detection of K-ras mutations on a DNA array. (a) Schematic representation of gel-based zip-code array. Glass microscope slides treated with g-methacryloyloxypropyltrimethoxysilane are used as the substrate for the covalent attachment of an acrylamide/acrylic acid copolymer matrix. Amine-modifed zip-code oligonucleotides are coupled to N-hydroxysuccinimide-activated surfaces at discrete locations (see Materials and Methods). Each position in the 3 * 3 grid identifes an individual zip-code address (and corresponding K-ras mutation or wild-type sequence). (b) Each robotically spotted array was hybridized with an individual LDR and fuorescent signal detected . All nine arrays identifed the correct mutant and/or wild-type for each tumor (G12S, G12R, and G12C) or cell line sample (Wt, G12D, G12A, G12V, and G13D). The small spots seen in some of the panels, e.g. near the center of the panel containing the G13D mutant, are not incorrect hybridizations, but noise due to imperfections in the polymer from Gerry et al., (1999). |
|
|
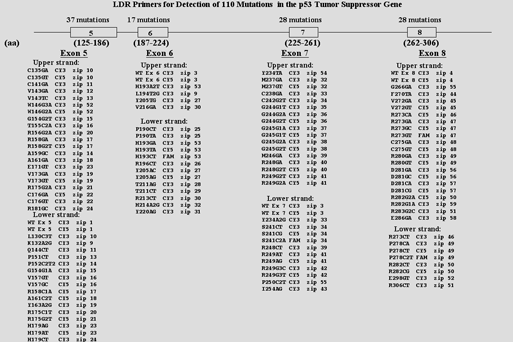
| Figure 7 and 8: Upper panel shows 216 LDR oligonucleotides required for detection of p53 mutation. Lower panel shows chip results. The mutation status of each sample and the zip-codes expected to capture signal are indicated at the bottom of each array; fiducials are along top and right side of all arrays. Two reactions were performed for each sample containing LDR primers that were designed to hybridize to the upper strand or lower strand of p53 sequence. The array was imaged on a Lumonics ScanArray 5000 to visualize the Cy3, Cy5 |
|
|
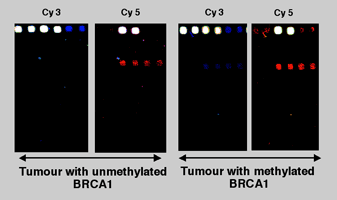
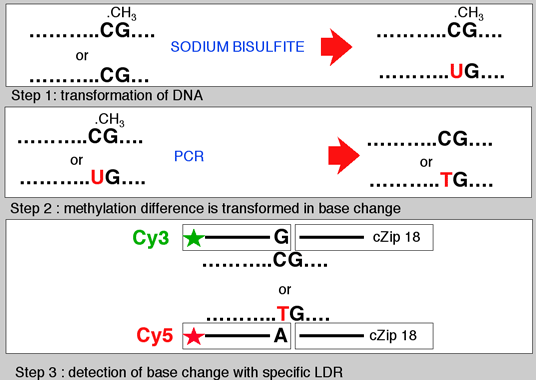
Figure 9 and 10: We have developed a sodium bisulfite-ligation-DNA array approach (Bisulfite-PCR/LDR/DNA array)to detect the presence or absence of DNA methylation at CpG sites within the promoters of the tumor suppressor genes, p15INK4b, p16Ink4a , p19ARF, p21Cip, p27Kip, p53, and BRCA1, as well as the imprinted gene SNRPN. |
References
Barany F and Gelfand DH (1991) Cloning, overexpression and nucleotide sequence of a thermostable DNA ligase-encoding gene. Gene 109: 1-11.
Barany F (1991) The ligase chain reaction in a PCR world. PCR Methods Appl 1: 5-16.
Barany F (1991) Genetic disease detection and DNA amplification using cloned thermostable ligase. Proc Natl Acad Sci U S A 88: 189-93.
Zebala JA and Barany F (1993) Implications for the ligase chain reaction in gastroenterology. J Clin Gastroenterol 17: 171-5.
Wiedmann M, Wilson WJ, Czajka J, Luo J, Barany F and Batt CA (1994) Ligase chain reaction (LCR)--overview and applications. PCR Methods Appl 3: S51-64.
Luo J, Bergstrom DE and Barany F (1996) Improving the fidelity of Thermus thermophilus DNA ligase. Nucleic Acids Res 24: 3071-8.
Zirvi M, Bergstrom DE, Saurage AS, Hammer RP and Barany F (1999) Improved fidelity of thermostable ligases for detection of microsatellite repeat sequences using nucleoside analogs. Nucleic Acids Res 27: e41.
Zirvi M, Nakayama T, Newman G, McCaffrey T, Paty P and Barany F (1999) Ligase-based detection of mononucleotide repeat sequences. Nucleic Acids Res 27: e40.
Gerry NP, Witowski NE, Day J, Hammer RP, Barany G and Barany F (1999) Universal DNA microarray method for multiplex detection of low abundance point mutations. J Mol Biol 292: 251-62.
Khanna M, Cao W, Zirvi M, Paty P and Barany F (1999) Ligase detection reaction for identification of low abundance mutations. Clin Biochem 32: 287-90.
Khanna M, Park P, Zirvi M, Cao W, Picon A, Day J, Paty P and Barany F (1999) Multiplex PCR/LDR for detection of K-ras mutations in primary colon tumors. Oncogene 18: 27-38.
Tong J, Cao W and Barany F (1999) Biochemical properties of a high fidelity DNA ligase from Thermus species AK16D. Nucleic Acids Res 27: 788-94.
Favis R and Barany F (2000) Mutation detection in K-ras, BRCA1, BRCA2, and p53 using PCR/LDR and a universal DNA microarray. Ann N Y Acad Sci 906: 39-43.
Favis R, Day JP, Gerry NP, Phelan C, Narod S and Barany F (2000) Universal DNA array detection of small insertions and deletions in BRCA1 and BRCA2. Nat Biotechnol 18: 561-4.
|
|